


WGBS+RNA-seq(項目文章)
Nature Communications | 華南師范大學任充華團隊通過WGBS揭示DNA甲基化在小蠊卵子發育的關鍵作用WGBS+RNA-seq(項目文章)
Nature Communications | 華南師范大學任充華團隊通過WGBS揭示DNA甲基化在小蠊卵子發育的關鍵作用表觀遺傳DNA甲基化提供了額外的基因組調控層面,增強了生物體的發育可塑性。5-甲基胞嘧啶(5mC)是動物中最常見和最豐富的DNA修飾。DNA甲基化過程由DNMT3負責重頭催化,隨后由DNMT1負責維持。通過調控基因表達,5mC修飾在調控基因組印跡、X染色體失活、染色體穩定性和染色質重塑等基本過程中起著關鍵作用。在大多數昆蟲目中,5mC修飾在不同程度上廣泛存在,并有助于種姓分化、長壽、繁殖和胚胎發生。然而,關于5mC修飾與昆蟲發育之間的機制聯系的知識有限,特別是在轉錄調控方面。
近日,華南師范大學生命科學學院昆蟲科學與技術研究所任充華研究員和李勝教授在Nature communications雜志發表題為“ 5mC modification orchestrates choriogenesis and fertilization by preventing prolonged ftz-f1 expression ” 的研究論文,該研究采用RNAi的技術手段,通過WGBS和RNA-seq的聯合分析,揭示了Dnmt1介導的5mC維持調控德國小蠊雌性繁殖的分子機制。艾斯基因為本研究提供了WGBS全基因組甲基化測序等技術服務。
英文標題: 5mC modification orchestrates choriogenesis and fertilization by preventing prolonged ftz-f1 expression
中文標題:5mC修飾通過抑制延長ftz-f1表達來協調絨毛膜的發生和受精
發表時間:2023年11月23日
發表期刊: Nature communications(IF = 16.6 / SCI 一區)
技術方案: RNA-seq、WGBS、NGS-BSP 、RNAi等
樣本類型: 德國小蠊雌性成蟲的六個組織部位(頭、腿、中腸、卵巢、胸腔和被膜)、整個第一生殖期(8天)的卵巢、dsCK和dsDnmt1處理后的卵巢等。
研究摘要
5-甲基胞嘧啶(5mC)的DNA甲基化是調控基因表達的重要表觀遺傳修飾。然而,迄今為止,對5mC如何調控昆蟲基因表達知之甚少。本研究揭示了Dnmt1介導的5mC維持調控德國小蠊雌性繁殖的分子機制。研究結果表明,Dnmt1的敲除降低了卵巢中5mC的水平,上調了絨毛膜發生過程中的許多基因,尤其是轉錄因子ftz-f1。在卵巢卵泡細胞中,ftz-f1啟動子區域的低甲基化增加并延長了ftz-f1的表達,從而導致20-羥基蛻皮激素(20E)水平異常升高和基質金屬蛋白酶(Mmp1) 基因的表達顯著上調。這些變化通過引起卵泡細胞的異常、絨毛膜蛋白的缺乏和海綿體的畸形而進一步損害絨毛膜的形成和破壞受精。這項研究極大地促進了對DNA 5mC修飾如何調節昆蟲雌性生殖的理解。
研究內容
(1)dnmt1介導的5mC維持是染色體發生和受精所必需的
為了研究德國小蠊5mC修飾與雌性生殖的關系,研究人員首先分析了德國小蠊雌性成蟲體內5mC修飾的時空分布。在6個組織中,與頭、腿和中腸相比,卵巢、胸腔和被膜中的5mC含量更高。隨后,研究人員還分析了整個第一生殖期(8天)卵巢內5mC的分布。結果表明,從第1天到第4天(成蟲羽化期(PAE))呈下降趨勢,隨后增加,直到產卵。qRT-PCR結果表明,第一個性腺周期卵巢中Dnmt1的表達水平與基因組5mC趨勢一致,即 DNMT1誘導的卵巢5mC維持可能在雌性生殖中起關鍵作用。
研究人員進一步在雌性成蟲中RNA干擾(RNAi)實驗,dsRNA被設計為靶向Dnmt1。dsDnmt1處理后基因表達顯著降低,5mC水平也顯著降低。dsDnmt1組卵囊全部萎縮、干癟,卵囊內的卵全部不能孵化。原核染色的結果表明,在ds CK處理的卵子中發現了來自卵子的雌性原核和來自精子的雄性原核,而在ds Dnmt1處理或未交配的卵子中未檢測到雄性原核。在dsDnmt1處理過的昆蟲中,海綿體嚴重畸形。這些結果表明dsDnmt1誘導的海綿體畸形破壞了受精,并進一步終止了胚胎發生。在dsDnmt1處理后的第7天(卵黃形成階段),卵巢卵泡細胞沒有表現出任何明顯的影響。然而,在PAE第9天(絨毛膜形成階段),dsDnmt1處理的卵巢中卵泡細胞核顯示不規則和凝聚的形狀,這些卵泡細胞顯示細胞骨架解體的跡象。這些發現表明dnmt1介導的5mC維持對德國蜚蠊的及時絨毛膜形成和正常受精至關重要。
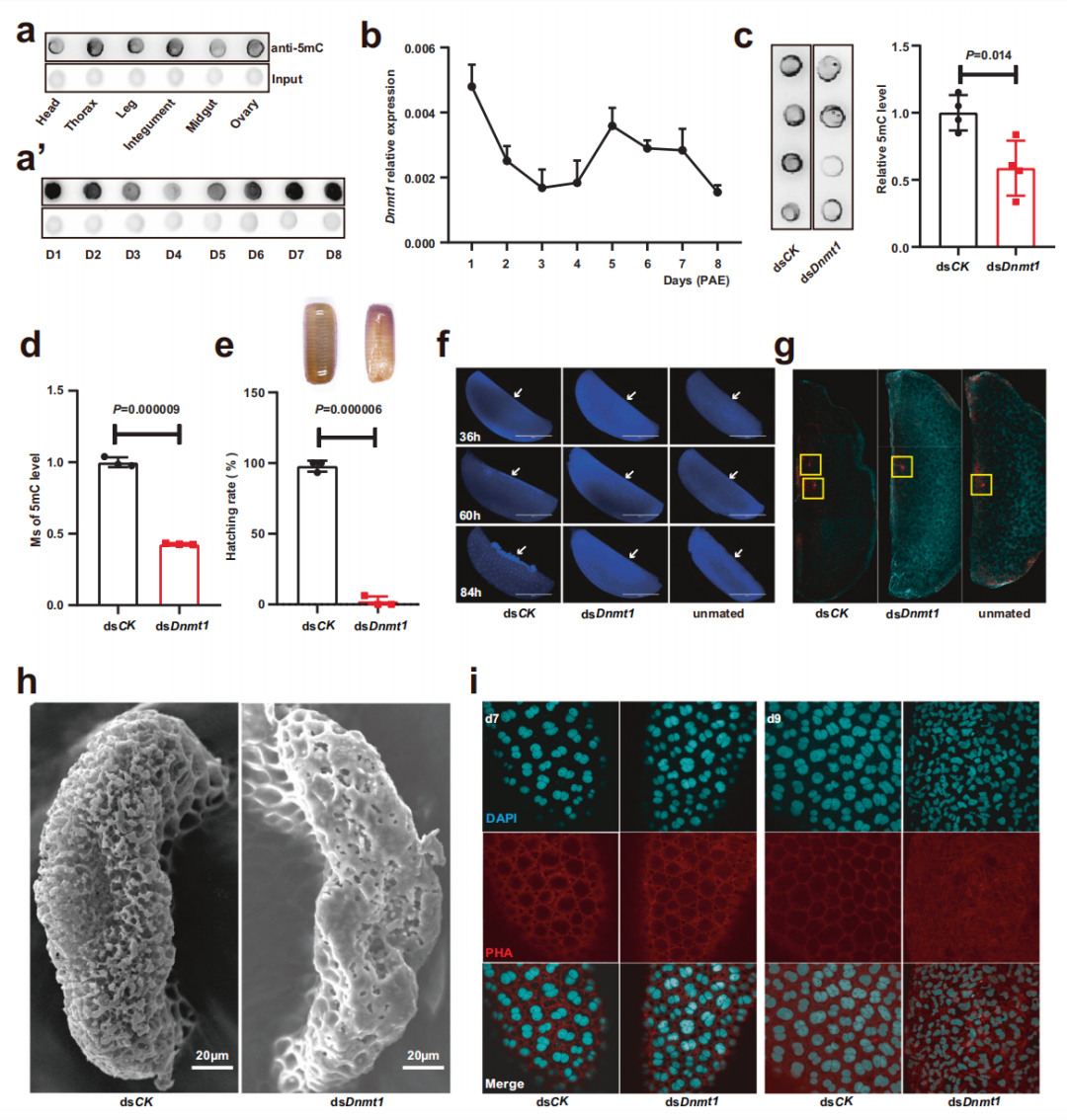
圖1 Dnmt1介導的5mC維持是絨毛膜形成和受精不可缺少的
(2)低甲基化誘導的過量20E損害絨毛膜的發生和受精
考慮到dsDnmt1在PAE第7天和第9天誘導的不同現象,研究人員在這兩個時間點對dsCK和dsDnmt1的卵巢進行了RNA-Seq檢測。主成分分析(PCA)和差異基因火山圖都表明,在dsCK組中,第7天和第9天的轉錄組有顯著差異;然而,dsDnmt1組在第7天和第9天的轉錄組與dsCK組在第7天的轉錄組非常相似。這些轉錄組學變化與兩個時間點觀察到的形態學變化一致,即dsDnmt1處理在第7天沒有顯著影響,但延遲了第9天的發育進展。
為了研究dsDnmt1對絨毛膜發生的影響,作者分析了PAE第9天上調的DEG,并檢測了它們在KEGG通路中的富集情況。幾種營養相關通路和昆蟲激素生物合成通路被激活和富集,這意味著營養或/和激素可能參與了DNMT1調控的絨毛膜發生。卵巢被報道是成年女性激素生成和循環20E的主要來源。研究人員觀察到dsDnmt1組在PAE第9天卵巢中20E水平增加了3倍。為了證實20E對絨毛膜發生的影響,作者將低劑量的20E注射到雌性成蟲體內,結果表明,大多數經20E處理的卵囊萎縮和干枯,其中大部分卵未能孵化。總之,dsDnmt1在絨毛膜形成和受精過程中誘導的表型缺陷應部分歸因于絨毛膜形成過程中20E水平的異常升高。
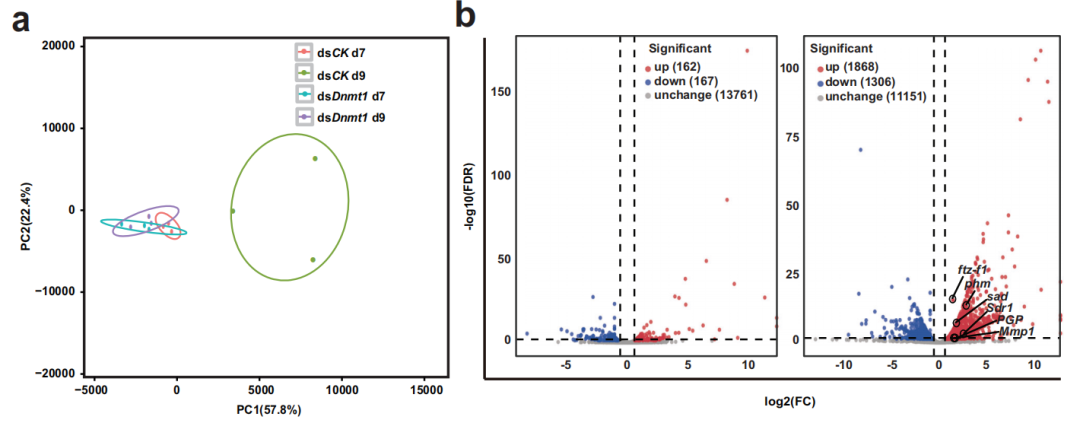
圖2低甲基化誘導的過量20E損害絨毛膜的發生和受精
(3)低甲基化增加和延長的ftz-f1表達會導致過度的類固醇生成
為了揭示差異基因表達與5mC修飾之間的聯系,研究人員利用WGBS,分析了dsDnmt1組和dsCK組在PAE第9天卵巢中5mC水平的差異。在dsDnmt1處理過的昆蟲卵巢中,CpG位點的全基因組甲基化水平從15%下降到6%。與dsCK組相比,dsDnmt1組中99.97%的差異甲基化區(DMRs)為低甲基化DMRs,僅發現0.03%的高甲基化DMRs。
隨后,作者探究了dsDnmt1處理昆蟲在PAE第9天卵巢中甾體生成變化與5mC水平變化之間的關系。作者發現,在dsDnmt1組中直接參與類固醇生成的DEGs中,ftz-f1顯著上調,并且上游 2kb啟動子區域顯著發生低甲基化,即Dnmt1可能通過維持ftz-f1啟動子區域的5mC水平來抑制甾體生成。研究人員進一步對dsDnmt1/dsCK中413個低甲基化和上調基因進行了分析,篩選出8個轉錄因子(TF)基因。然后分別敲除前6個TF基因,發現只有dsftz-f1處理顯著降低了phm(phantom)的表達。敲除ftz-f1也導致EcR表達顯著降低。此外,敲除phm、ftz-f1、EcR或taiman (tai, EcR的共激活劑)可顯著降低20E水平。這些發現表明,FTZ-F1可能不僅作為促進甾體生成的轉錄因子,而且作為與Tai一起增強20E信號通路EcR活性的能力因子。總之,作者得出結論,dsDnmt1引起的低甲基化增加并延長了ftz-f1的表達,導致過度的類固醇生成。
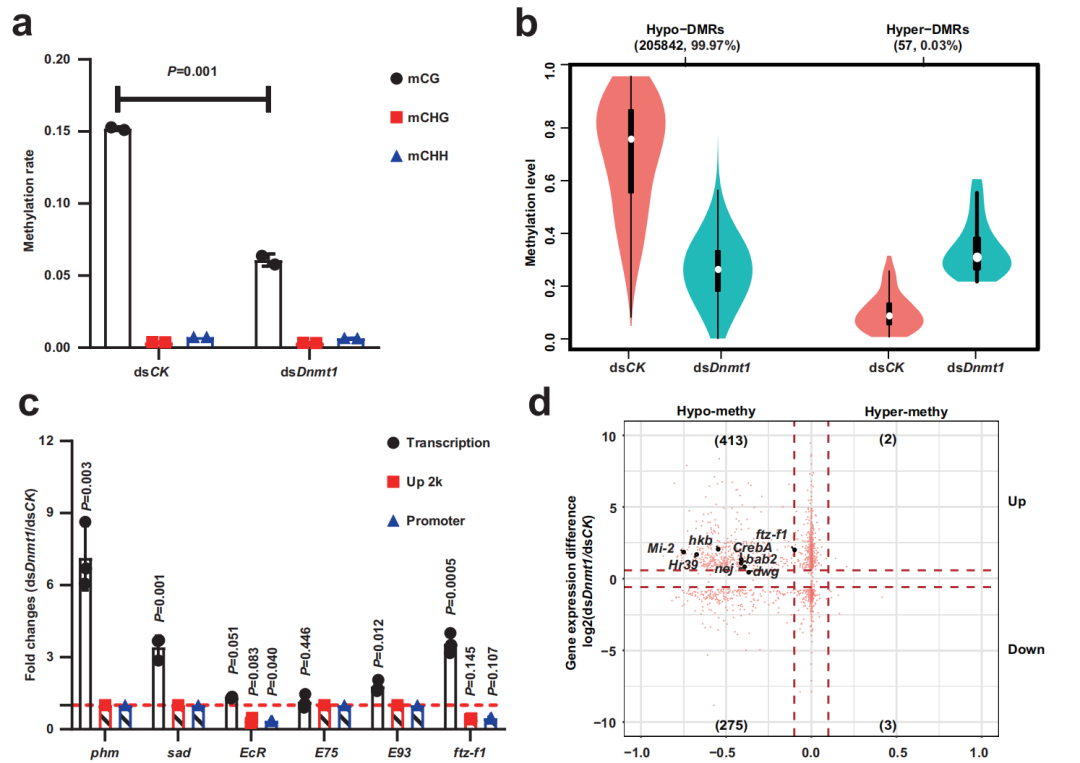
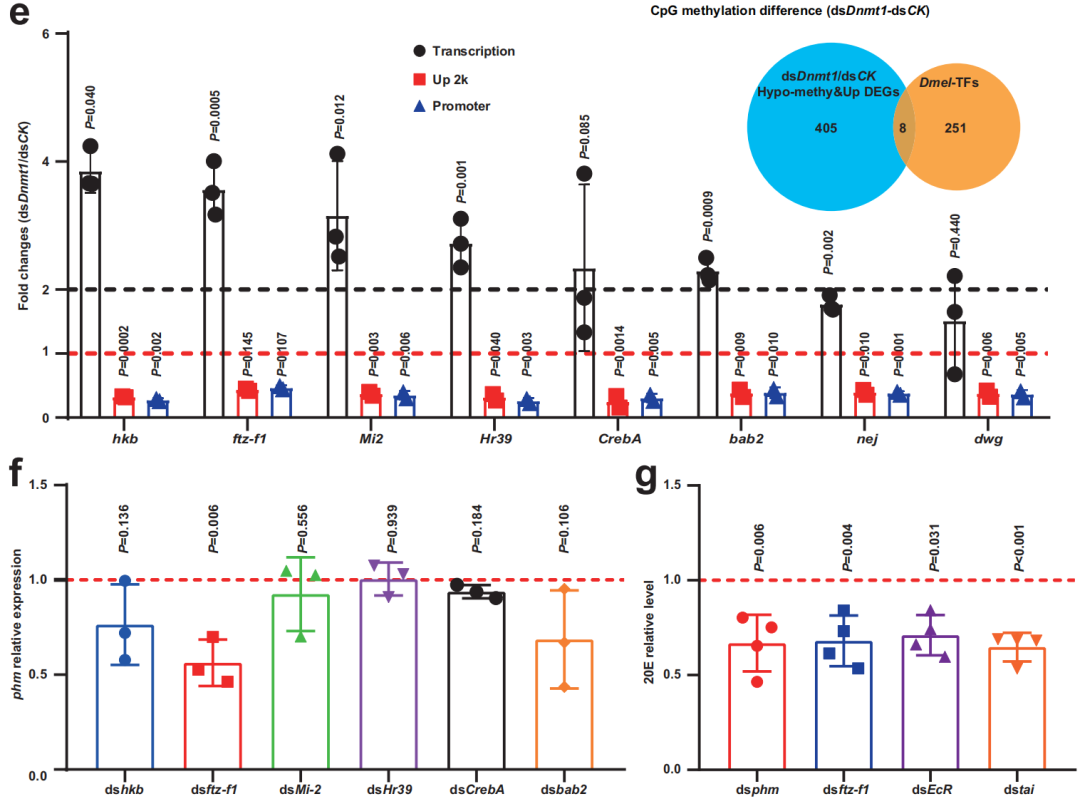
圖3低甲基化誘導的ftz-f1過表達調節卵泡細胞的卵狀細胞形成
(4) ftz-f1啟動子通過5mC修飾以防止ftz-f1的表達
研究人員進一步研究了dsDnmt1誘導的低甲基化與第9天PAE時ftz-f1表達延長之間的潛在聯系。WGBS數據表明,在dsDnmt1組中觀察到ftz-f1啟動子區域中5mC水平急劇下降。此外,下一代亞硫酸鹽測序PCR (NGS-BSP)的結果也表明,在ftz-f1啟動子區域的6個擴增子(~7kb)中觀察到5mC水平急劇下降。作者進一步使用NGS-BSP和RNA-seq評估了第一個性腺周期中ftz-f1啟動子中5mC和表達水平的變化趨勢。對相同的6個擴增子進行分析,從PAE第1-3天到PAE第4-6天,擴增子3和5有明顯的下降趨勢,隨后增加直到產卵。并且,ftz-f1的表達一直保持在低水平直到PAE第6天,在卵黃發生結束時達到峰值(第7天),然后在絨毛膜發生期間下降。總的來說,ftz-f1這兩個啟動子區域的5mC水平與ftz-f1表達水平呈負相關,表明ftz-f1啟動子通過Dnmt1介導的5mC修飾來阻止ftz-f1表達的延長。
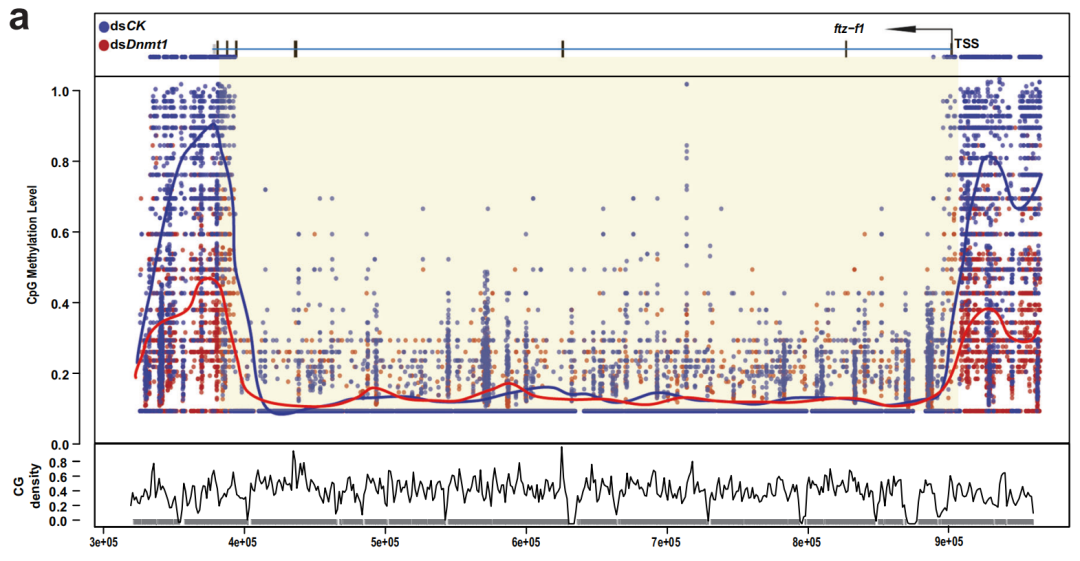
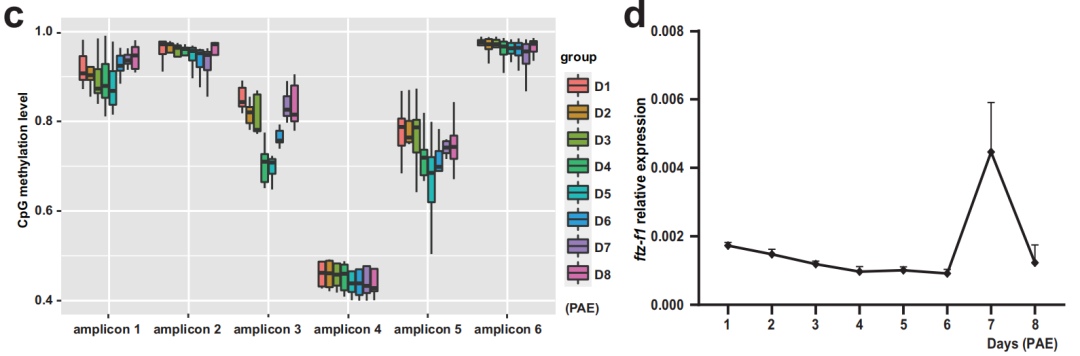
圖4 ftz-f1啟動子通過5mC修飾抑制ftz-f1延長表達
(5)dsDnmt1誘導的缺陷部分被ftz-f1挽救
為了證實Dnmt1對ftz-f1表達的負調控作用,研究人員同時進行了dsDnmt1和dsftz-f1處理。與dsftz-f1共處理可減弱dsnmt1誘導的ftz-f1上調,dsDnmt1誘導的過量20E水平也被dsftz-f1的共同處理部分抵消。由于dsftz-f1處理可以完全破壞產卵過程,因此與dsDnmt1共處理使產卵率恢復到較高水平。此外,作者還關注了濾泡細胞和海綿體上明顯的表型缺陷。dsDnmt1與dsftz-f1共處理部分挽救了dsDnmt1誘導的卵泡細胞解體和海綿體畸形形成。dsDnmt1和dsftz-f1共處理的結果進一步表明,dnmt1在ftz-f1啟動子區域維持5mC水平抑制了ftz-f1的表達,dsDnmt1-增加和延長的ftz-f1表達破壞了絨毛膜的發生和受精。
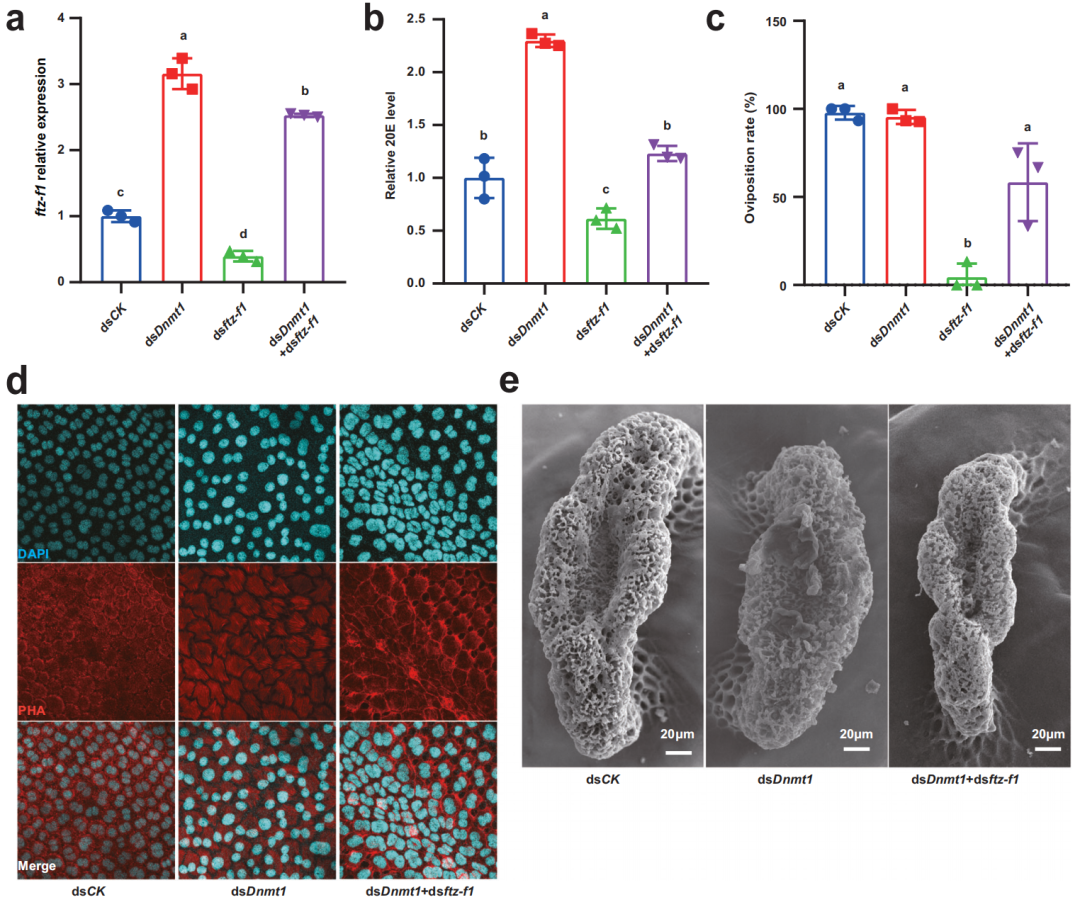
圖5在絨毛膜發生過程中,ftz-f1基因的敲除可以部分挽救dsDnmt1誘導的現象
(6)5mC修飾可阻止ftz-f1誘導的Mmp1表達和突變
已有研究表明,FTZ-F1誘導基質金屬蛋白酶 (Mmp)基因的表達,從而調控果蠅卵巢卵泡細胞的成熟和破裂。與這一調控機制一致,Mmp1在dsDnmt1處理的卵巢中上調,并被dsDnmt1和dsftz-f1雙RNAi挽救。當ftz-f1被敲除時,卵巢中Mmp1的表達也顯著下調。此外,作者還研究了5個已知的絨毛膜基因在轉錄組中的表達, 其中3個在dsDnmt1組的PAE第9天顯著下調。總的來說,低甲基化增加和延長的ftz-f1表達導致卵泡細胞解離和失活,這可能導致無法合成足夠的絨毛膜蛋白,進一步破壞受精。
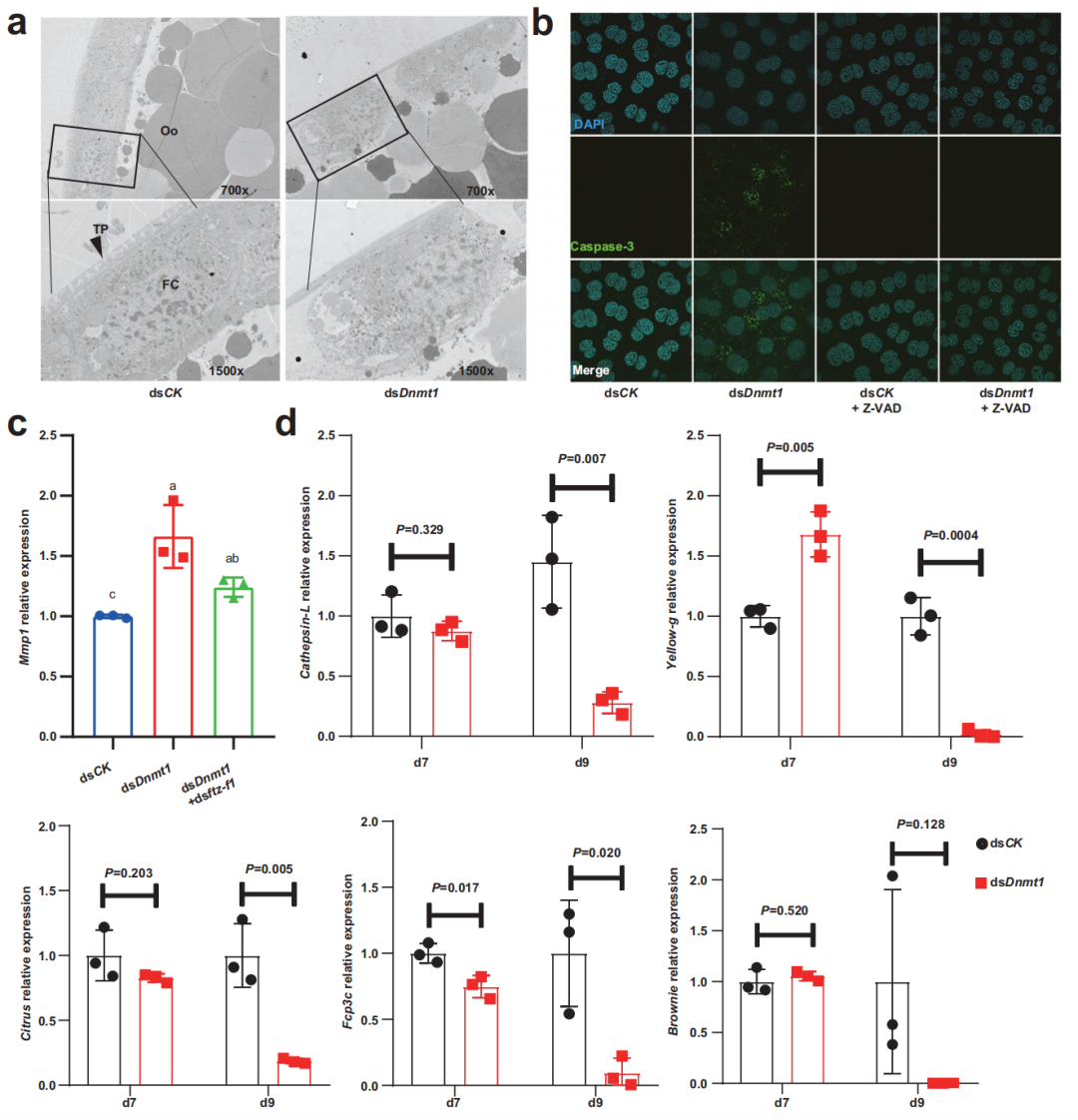
圖6 5mC修飾可防止絨毛發生過程中ftz-f1誘導的失活
研究小結
綜上所述,本研究首次證明了昆蟲中基因啟動子區域的DNA甲基化與基因的轉錄水平呈負相關性,并找到了串聯DNA 5mC修飾和卵子發育的關鍵“開關”基因ftz-f1,為昆蟲繁殖生物學研究提供了新的見解,也為害蟲防治提供了新的思路和分子靶標。
www.vgrsoft.com在線搜索取消
清空記錄
歷史記錄
清空記錄
歷史記錄
深圳市艾斯基因科技有限公司專注表觀組學技術創新及臨床醫學轉化應用15年
產品中心深圳市艾斯基因科技有限公司關注我們 粵ICP備16072881號-3 粵ICP備16072881號-4版權所有 ? 深圳市艾斯基因科技有限公司粵ICP備16072881號-1
粵ICP備16072881號-3 粵ICP備16072881號-4版權所有 ? 深圳市艾斯基因科技有限公司粵ICP備16072881號-1 選擇區號
選擇區號
 瀏覽器自帶分享功能也很好用哦~?
瀏覽器自帶分享功能也很好用哦~?復制成功
×
 瀏覽器自帶分享功能也很好用哦~
瀏覽器自帶分享功能也很好用哦~